Термин ретикулярная формация предложил в 1865 году немецкий ученый О. Дейтерс. Под этим термином Дейтерс понимал разбросанные в стволе головного мозга клетки, окруженные множеством волокон, идущих в различных направлениях. Именно сетевидное расположение волокон, связывающих между собой нервные клетки, послужило основой для предложенного названия.
В настоящее время морфологами и физиологами накоплен богатый материал о строении и функциях ретикулярной формации. Установлено, что структурные элементы ретикулярной формации локализуются в целом ряде мозговых образований, начиная с промежуточной зоны шейных сегментов спинного мозга (VII пластина), и заканчивая некоторыми структурами промежуточного мозга (интраламинарными ядрами, таламическим ретикулярным ядром). Ретикулярная формация состоит из значительного числа нервных клеток (в ней содержится почти 9/10 клеток всего ствола мозга). Общие черты строения ретикулярных структур - наличие особых ретикулярных нейронов и отличительный характер связей.
Рис. 1. Нейрон ретикулярной формации. Сагиттальный разрез ствола мозга крысенка.
На рисунке А представлен только один нейрон ретикулярной формации. Видно, что аксон разделяется на каудальный и ростральный сегменты, большой протяженности, со множеством коллатералей. Б. Коллатерали. Сагиттальный разрез нижней части ствола мозга крысенка, показывающий соединения коллатералей большого нисходящего пути (пирамидный путь) с ретикулярными нейронами. Коллатерали восходящих путей (сенсорные пути), отсутствующие на рисунке, соединяются с ретикулярными нейронами подобным же образом (по Шейбэлу М. Э. и Шейбэлу А. Б.)
Наряду с многочисленными отдельно лежащими нейронами, различными но форме и величине, в ретикулярной формации головного мозга имеются ядра. Рассеянные нейроны ретикулярной формации прежде всего играют важную роль в обеспечении сегментарных рефлексов, замыкающихся на уровне ствола головного мозга. Они выступают в качестве вставочных нейронов при осуществлении таких рефлекторных актов, как моргание, роговичный рефлекс и т. д.
Выяснено значение многих ядер ретикулярной формации. Так, ядра, расположенные в продолговатом мозге, имеют связи с вегетативными ядрами блуждающего и языкоглоточного нервов, симпатическими ядрами спинного мозга, они участвуют в регуляции сердечной деятельности, дыхания, тонуса сосудов, секреции желез и т. д.
Установлена роль голубого пятна и ядер шва в регуляции сна и бодрствования. Голубое пятно , находится в верхнелатеральной части ромбовидной ямки. Нейроны этого ядра продуцируют биологически активное вещество - норадреналин , который оказывает активирующее воздействие на нейроны вышележащих отделов мозга. Особенно высока активность нейронов голубого пятна во время бодрствования, во время глубокого сна она угасает почти полностью. Ядра шва располагаются по срединной линии продолговатого мозга. Нейроны этих ядер вырабатывают серотонин , который вызывает процессы разлитого торможения и состояние сна.
Ядра Кахаля и Даркшевича , относящиеся к ретикулярной формации среднего мозга, имеют связи, с ядрами III, IV, VI, VIII и XI пар черепных нервов. Они координируют работу этих нервных центров, что очень важно для обеспечения сочетанного поворота головы и глаз. Ретикулярная формация ствола головного мозга имеет важное значение в поддержании тонуса скелетной мускулатуры, посылая тонические импульсы на мотонейроны двигательных ядер черепных нервов и двигательных ядер передних рогов спинного мозга. В процессе эволюции из ретикулярной формации выделились такие самостоятельные образования, как красное ядро, черное вещество.
По структурно-функциональным критериям ретикулярная формация делится на 3 зоны:
1. Медианную, расположенную по средней линии;
2. Медиальную, занимающую медиальные отделы ствола;
3. Латеральную, нейроны которой лежат вблизи сенсорных образований.
Медианная зона представлена элементами шва, состоящие из ядер, нейроны которых синтезируют медиатор – серотонин. Система ядер шва принимает участие в организации агрессивного и полового поведения, в регуляции сна.
Медиальная (осевая) зона состоит из мелких нейронов, которые не ветвятся. В зоне располагается большое количество ядер. Встречаются также крупные мультиполярные нейроны с большим числом густо ветвящихся дендритов. Они образуют восходящие нервные волокна в кору больших полушарий и нисходящие нервные волокна в спинной мозг. Восходящие пути связи медиальной зоны оказывают активирующее влияние (прямо или опосредованно через таламус) на новую кору. Нисходящие пути оказывают тормозящее влияние.
Латеральная зона – к ней относятся ретикулярные образования, расположенные в стволе мозга вблизи сенсорных систем, а также ретикулярные нейроны, лежащие внутри сенсорных образований. Основным компонентом этой зоны является группы ядер, которые примыкают к ядру тройничного нерва. Все ядра латеральной зоны (за исключением ретикулярного латерального ядра продолговатого мозга) состоят из нейронов малой и средней величины и лишены крупных элементов. В этой зоне располагаются восходящие и нисходящие пути, обеспечивающие связь сенсорных образований с медиальной зоной ретикулярной формации и моторными ядрами ствола. Эта часть ретикулярной формации является более молодым и возможно прогрессивнее, с ее развитием связан факт уменьшения объема осевой ретикулярной формации в ходе эволюционного развития. Таким образом, латеральная зона – это совокупность элементарных интегративных единиц, сформированных вблизи и внутри специфических сенсорных систем.
Рис. 2. Ядра ретикулярной формации (РФ) (по: Niuwenhuys еt. аl, 1978).
 |
1-6 - медианная зона РФ: 1-4- ядра шва (1 - бледное, 2 - темное, 3 - большое, 4- мостовое), 5 - верхнее центральное, 6 - дорсальное ядро шва, 7-13 - медиальная зона РФ: 7 - ретикулярное парамедианное, 8 - гигантоклеточное, 9 - ретикулярное ядро покрышки моста, 10, 11 - каудальное (10) и оральное (11) ядра моста, 12 - дорсальное покрышечное ядро (Гуддена), 13 - клиновидное ядро, 14- I5-латеральная зона РФ: 14 - центральное ретикулярное ядро продолговатого мозга, 15 - латеральное ретикулярное ядро, 16, 17 - медиальное (16) и латеральное (17) парабрахиальные ядра, 18, 19 - компактная (18) и рассеянная (19) части педункуло-понтийного ядра.
Благодаря нисходящим влияниям ретикулярная формация оказывает тоническое влияние и на мотонейроны спинного мозга, что в свою очередь повышает тонус скелетной мускулатуры, совершенствует систему обратной афферентной связи. В результате любой двигательный акт совершается значительно эффективнее, осуществляет более точный контроль за движением, но чрезмерное возбуждение клеток ретикулярной формации может привести к дрожанию мышц.
В ядрах ретикулярной формации находятся центры сна и бодрствования, и стимуляция тех или иных центров приводит или к наступлению сна, или к пробуждению. На этом основано применение снотворных. В ретикулярной формации расположены нейроны, реагирующие на болевые раздражения, идущие от мышц или внутренних органов. В ней также расположены специальные нейроны, которые обеспечивают быструю реакцию на внезапные, неопределенные сигналы.
Ретикулярная формация тесно связана с корой больших полушарий, благодаря этому формируется функциональная связь между внешними отделами ЦНС и стволом головного мозга. Ретикулярная формация играет важную роль как в интеграции сенсорной информации, так и в контроле над деятельностью всех эффекторных нейронов (моторных и вегетативных). Она имеет также первостепенное значение для активации коры больших полушарий, для поддержания сознания.
Необходимо отметить, что кора полушарий большого мозга, и в свою очередь, посылает по корково-ретикулярным путям импульсы в ретикулярную формацию. Эти импульсы возникают в основном в коре лобной доли и проходят в составе пирамидных путей. Корково-ретикулярные связи оказывают либо тормозное, либо возбуждающее действие на ретикулярную формацию ствола головного мозга, они осуществляют корректировку прохождения импульсов по эфферентным путям (отбор эфферентной информации).
Таким образом, между ретикулярной формацией и корой полушарий большого мозга имеется двусторонняя связь, которая обеспечивает саморегуляцию в деятельности нервной системы. От функционального состояния ретикулярной формации зависит тонус мускулатуры, работа внутренних органов, настроение, концентрация внимания, память и т. д. В целом ретикулярная формация создает и поддерживает условия для осуществления сложной рефлекторной деятельности с участием коры полушарий большого мозга.
Ретикулярная (сетчатая) формация представляет собой скопления нейронов, различных по функции и размерам, связанных множеством нервных волокон, проходящих в разных направлениях и образующих сеть на всем протяжении ствола мозга, что и определяет ее название. Нейроны расположены либо диффузно, либо образуют ядра.
Нейроны ретикулярной формации имеют длинные маловетвящиеся дендриты и хорошо ветвящиеся аксоны, которые часто образуют Т-образное ветвление: одна из ветвей аксона имеет нисходящее, а вторая - восходящее направление.
А. Функциональные особенности нейронов ретикулярной формации:
подимодалыюстъ - для нейронов ретикулярной формации характерна полисенсорная конвергенция, они принимают коллатерали от нескольких сенсорных путей, идущих от разных рецепторов;
тоническая активность, в покое равная 5-10 имп/с;
высокая чувствительность к некоторым веществам крови (например, адреналину, СО2) и лекарствам (барбитуратам, аминазину и др.);
Более выраженная возбудимость по сравнению с другими нейронами;
высокая лабильность - до 500-1000 имп/с.
Нейроны и ядра ретикулярной формации входят в состав центров, регулирующих функции внутренних органов (кровообращения, дыхания, пищеварения), тонус скелетной мускулатуры (см. раздел 5.3), активность коры большого мозга. Обширны связи ретикулярной формации с другими отделами ЦНС и рефлексогенными зонами: она получает импульсацию от различных рецепторов организма и отделов ЦНС и в свою очередь посылает импульсы во все отделы мозга. При этом выделяют восходящие и нисходящие влияния ретикулярной формации.
Б. Нисходящие влияния ретикулярной формации на моторные спинальные центры. От ретикулярного гигантоклеточного ядра продолговатого мозга идет частично перекрещенный латеральный ретикулоспинальный тракт, волокна которого оканчиваются на вставочных нейронах спинного мозга. Через эти интернейроны они возбуждают а- и у-мотонейроны мышц-сгибателей мускулатуры конечностей и реципрокно тормозят с помощью тормозных интернейронов мышцы-разгибатели.
От каудальных и оральных ретикулярных ядер моста идет неперекрещенный медиальный ретикулоспинальный тракт, волокна которого оканчиваются на интернейронах спинного мозга. Через них осуществляется стимуляция а- и у-мотонейронов мышц-разгибателей, а через тормозные интернейроны тормозятся мышцы-сгибатели. О роли ретикулярной формации моста, продолговатого мозга в регуляции тонуса мышц-разгибателей см. рис. 5.9.
В. Восходящие влияния РФ на большой мозг могут быть как активирующими, так и тормозными. Импульсы ретикулярных нейронов продолговатого мозга (гигантоклеточное, латеральное и вентральное ретикулярные ядра), моста (особенно каудальное ретикулярное ядро) и среднего мозга поступают к неспецифическим ядрам таламуса, и после переключения в них проецируются в различные области коры. Кроме таламуса восходящие влияния поступают также в задний гипоталамус, полосатое тело.
В эксперименте после перерезки ствола мозга между верхними и нижними холмиками четверохолмия (изолированный передний мозг) у животного не нарушалось поступление в кору большого мозга возбуждения по важнейшим сенсорным системам - зрительной и обонятельной. Однако животное вело себя как спящее: у него был нарушен контакт с внешним миром, оно не реагировало на световые и обонятельные раздражители (спящий мозг по Бремеру). На ЭЭГ у таких животных преобладали медленноволновые регулярные ритмы. У человека аналогичные ритмы возникают при спокойном бодрствовании и в дремотном состоянии. Такое же состояние головного мозга (спящий мозг) наблюдается при повреждении только восходящих путей ретикулярной формации.
Стимуляция ретикулярной формации вызывает пробуждение животного [Мегун Г., Моруцци Дж., 1949]. На ЭЭГ медленные ритмы сменяются при этом высокочастотными ритмами (реакция десинхронизации), свидетельствующими об активированном состоянии коры головного мозга. На основании полученных данных сложилось представление о том, что важнейшей функцией восходящей ретикулярной формации является регуляция цикла сон/бодрствование и уровня сознания.
Тормозное влияние ретикулярной формации на большой мозг изучено значительно хуже. В.Гесс (1929), Дж.Моруцци (1941) установили, что раздражением некоторых точек ретикулярной формации ствола мозга можно перевести животное из бодрствующего состояния в сонное. При этом возникает реакция синхронизации ритмов ЭЭГ.
СИСТЕМЫ СВЯЗЕЙ СТВОЛА МОЗГА
Связи различных отделов ЦНС осуществляются с помощью нервных путей, идущих в различных направлениях и выполняющих разные функции, что и положено в основу их классификации. В частности, в спинном мозге, как и в других отделах ЦНС, выделяют восходящие и нисходящие пути (определяющим фактором этой классификации является направление потока импульсов).
Кроме того, в стволе мозга восходящие системы подразделяют на специфические и неспецифические.
Восходящие и нисходящие пути спинного мозга рассмотрены в разделе 5.2.2.
Проводниковая функция ствола мозга выполняется нисходящими и восходящими путями, часть из которых переключается в стволовых центрах, другая часть идет транзиторно (без переключения).
А. Восходящие пути являются частью проводникового отдела анализаторов, передающих информацию от рецепторов в проекционные зоны коры. В стволе мозга выделяют две восходящие системы: специфическую и неспецифическую.
1. Специфическую восходящую систему составляет лемнискота-ламический путь, в котором выделяют медиальную и латеральную петли. Медиальная петля образуется преимущественно из аксонов нейронов тонкого ядра (Голля) и клиновидного ядра (Бурдаха), которые проводят импульсы от проприорецепторов. Волокна медиальной петли переключаются в вентральных задних специфических ядрах таламуса. Медиальная петля входит в проводниковый отдел слухового анализатора, ее волокна переключаются в медиальном коленчатом теле таламуса и нижних буграх четверохолмия. К специфической проводящей системе относятся проводящие пути зрительного и вестибулярного анализаторов. Импульсы по специфическим восходящим путям поступают в корковый конец соответствующего анализатора (зрительного, слухового и т.д.).
2. Неспецифические (экстралелтисковые) восходящие пути переключаются в неспецифических (интраламинарных и ретикулярном) ядрах таламуса. В основном это волокна латерального спиноталамического и спиноретикулярного трактов, проводящих температурную и болевую чувствительность. Импульсация от них проецируется в различные области коры (особенно лобную орбитальную кору). Неспецифическая система получает коллатеральные волокна от специфической системы, что обеспечивает связь этих двух восходящих систем. Функциональной особенностью неспецифической системы является относительно медленное проведение возбуждения. Рецептивные поля нейронов большие, нейроны гюлимодаль-ные, связанные с несколькими видами чувствительности, топография проекции периферии в центрах не выражена.
3. Часть афферентной импульсации поступает в мозжечок по другим системам. Через ствол мозга в мозжечок проходят задний спинно-мозжечковый тракт Флексига и передний спинно-мозжечковый тракт Говерса, проводящие импульсацию от рецепторов мышц и связок, а также вестибуломозжечковый тракт, несущий информацию от вестибулярных рецепторов. Из коры мозжечка информация передается в вентральные ядра таламуса, далее она проецируется в соматосенсорную, моторную и премоторные зоны коры большого мозга.Б. Нисходящие проводниковые пути ствола мозга включают двигательные пирамидные пути, начинающиеся от клеток Беца коры прецентральной извилины. Они иннервируют мотонейроны передних рогов спинного мозга (кортикоспинальные пути), мотонейроны двигательных ядер черепных нервов (кортикобульбарный путь), обеспечивая произвольные сокращения мышц конечностей, туловища, шеи и головы. Моторные центры ствола мозга и их пути - важнейший компонент экстрапирамидной системы, основной функцией которой является регуляция мышечного тонуса, позы и равновесия. К этой системе на уровне ствола мозга относятся кортикорубральный и кортикоретику-лярный тракты, оканчивающиеся на моторных центрах ствола, от которых идут рубро-, ретикуло- и вестибулоспинальный пути. Экстрапирамидная система представляет собой совокупность ядер ствола мозга внепирамидной системы. Ее основными элементами являются: полосатое тело, бледный шар, красное ядро, ретикулярная формация.
В стволе мозга проходят нисходящие пути, обеспечивающие двигательные функции мозжечка; к ним относится кортико-мостомозжечковый путь, по которому в мозжечок поступает импульсация от двигательной и других областей коры. Обработанная в коре мозжечка и его ядрах информация поступает на моторные ядра ствола (красное, вестибулярные, ретикулярные). Через ствол мозга проходит начинающийся в четверохолмии тектоспинальный тракт, который обеспечивает двигательные реакции организма в ориентировочных зрительных и слуховых рефлексах. Все двигательные реакции организма осуществляются нисходящими системами с помощью а- и у-мотонейронов спинного мозга и нейронов двигательных ядер черепных нервов.
МОЗЖЕЧОК
Мозжечок расположен позади полушарий большого мозга, над продолговатым мозгом и мостом. В совокупности с последним он образует задний мозг. Мозжечок включает в себя более половины всех нейронов ЦНС, хотя составляет 10% массы головного мозга. Это свидетельствует о больших возможностях обработки информации мозжечком. Он играет важную роль в интеграции двигательных и вегетативных реакций, в частности в координации произвольных и непроизвольных движений, поддержании равновесия, регуляции мышечного тонуса.
А. Функциональная организация. Выделяют три структуры мозжечка, отражающие эволюцию его функций:
Древний мозжечок (архицеребеллум) состоит из клочка и узелка (флоккулонодулярная доля) и нижней части червя; имеет наиболее выраженные связи с вестибулярной системой, поэтому его называют также вестибулярным мозжечком;
Старый мозжечок (палеоцеребеллум) включает верхнюю часть червя, парафлоккулярный отдел, пирамиды и язычок; получает информацию преимущественно от проприорецепторов. Его называют также спинальным мозжечком;
Новый мозжечок (неоцеребеллум) состоит из двух полушарий. Он получает информацию от коры, преимущественно по лобно-мостомозжечковому пути, от зрительных и слуховых ре-цептирующих систем, что свидетельствует об его участии в анализе зрительных, слуховых сигналов и организации на них реакции.
1. Межнейронные связи в коре полушарий мозжечка, его афферентные входы и эфферентные выходы весьма разнообразны. Грушевидные нейроны (клетки Пуркинье) образуют средний -II (ганглиозный) слой коры, являющейся главной функциональной единицей мозжечка. Структурной основой служат многочисленные ветвящиеся дендриты, на которых в одной клетке может быть до 100000 синапсов.
Клетки Пуркинье являются единственными эфферентными нейронами коры мозжечка и обеспечивают его связь с корой большого мозга, стволовыми образованиями и спинным мозгом. Эти клетки непосредственно связывают его кору с внутримозжечковыми и вестибулярными ядрами. В связи с этим функциональное влияние мозжечка существенным образом зависит от активности клеток Пуркинье.
Информация к клеткам Пуркинье (афферентные входы) поступает практически от всех рецепторов: мышечных, вестибулярных, кожных, зрительных, слуховых; от нейронов основания задних рогов спинного мозга (по спинно-оливному пути), а также от двигательной коры головного мозга, ассоциативной коры и ретикулярной формации.
На мозжечок передается влияние некоторых структур ствола головного мозга, например голубого пятна и ядер шва.
Преобладающее как прямое, так и опосредованное афферентное влияние на клетки Пуркинье является возбуждающим. Но поскольку клетки Пуркинье являются тормозными нейронами (медиатор ГАМК), то с их помощью кора мозжечка превращает возбуждающие сигналы на входе в тормозные сигналы на выходе. Таким образом, эфферентное влияние коры мозжечка на последующее нейронное звено (в основном это внутримозжечковые ядра) является тормозным. Под II слоем коры (под клетками Пуркинье) лежит гранулярный (III) слой, состоящий из клеток-зерен, число которых достигает 10 млрд. Аксоны этих клеток поднимаются вверх, Т-образно делятся на поверхности коры, образуя дорожки контактов с клетками Пуркинье. Здесь же лежат клетки Гольджи.
Верхний (I) слой коры мозжечка - молекулярный, состоит из параллельных волокон, разветвлений дендритов и аксонов II и III слоев. В нижней части молекулярного слоя встречаются корзинчатые и звездчатые клетки, которые обеспечивают взаимодействие клеток Пуркинье.
Стимуляция верхнего слоя коры мозжечка приводит к длительному (до 200 мс) торможению активности клеток Пуркинье. Такое же их торможение возникает при световых и звуковых сигналах. Суммарные изменения электрической активности коры мозжечка на раздражение чувствительного нерва любой мышцы вызывают торможение активности коры (гиперполяризация клеток Пуркинье), которое наступает через 15-20 мс и длится 20-30 мс, после чего возникает волна возбуждения, длящаяся до 500 мс (деполяризация клеток Пуркинье).
Фоновая импульсная активность нейронов регистрируется в слое клеток Пуркинье и гранулярном слое, причем частота генерации импульсов этих клеток колеблется от 20 до 200 в секунду.
2. Подкорковая система мозжечка включает три функционально разных ядерных образования: ядро шатра, пробковидное, шаровидное и зубчатое ядра.
Ядро шатра получает информацию от медиальной зоны коры мозжечка и связано с ядром Дейтерса и ретикулярной формацией продолговатого и среднего мозга. Отсюда сигналы идут по ретикулоспинальному пути к мотонейронам спинного мозга.
На пробковидное и шаровидное ядра проецируется промежуточная кора мозжечка. От них связи идут в средний мозг к красному ядру, далее в спинной мозг по руброспинальному пути.
Зубчатое ядро получает информацию от латеральной зоны коры мозжечка, оно связано с таламусом, а через него - с моторной зоной коры большого мозга.
Клетки ядер мозжечка значительно реже генерируют импульсы (1-3 в секунду), чем клетки коры мозжечка (клетки Пуркинье -20-200 импульсов в секунду).
3. С соседними отделами мозга мозжечок соединяется тремя парами ножек. Нижние мозжечковые ножки соединяют мозжечок с продолговатым мозгом, средние - с мостом, верхние - со средним мозгом. По проводящим путям ножек мозжечок получает афферентную импульсацию (входы) от других отделов мозга и посылает эфферентные импульсы (выходы) к различным структурам мозга.
Через верхние ножки сигналы идут в таламус, мост, красное ядро, ядра ствола мозга, в ретикулярную формацию среднего мозга. Средние ножки мозжечка связывают новый мозжечок с лобной долей мозга. Через нижние ножки мозжечка сигналы идут в продолговатый мозг, к его вестибулярным ядрам, оливам, ретикулярной формации.
Афферентная импульсация в кору мозжечка от кожных рецепторов, мышц, суставных оболочек, надкостницы поступает по так называемым спинно-мозжечковым трактам: заднему (дорсальному) и переднему (вентральному). Эти пути к мозжечку проходят через нижнюю оливу продолговатого мозга. От клеток олив идут так называемые лазающие волокна, которые ветвятся на дендритах клеток Пуркинье.
Ядра моста посылают афферентные пути в мозжечок, образующие мшистые волокна, которые оканчиваются на клетках-зернах III слоя коры мозжечка. Между мозжечком и голубым пятном среднего мозга существует афферентная связь с помощью адренергических волокон. Эти волокна способны диффузно выбрасывать норадреналин в межклеточное пространство коры мозжечка, тем самым гуморально изменяют состояние возбудимости его клеток.
Рассмотренная структурно-функциональная организация нейронов мозжечка позволяет понять соматические и вегетативные его функции.
Б, Двигательные функции мозжечка состоят в регуляции мышечного тонуса, позы и равновесия, координации выполняемого целенаправленного движения, программировании целенаправленных движений.
1. Мышечный тонус и поза регулируются преимущественно древним мозжечком (флоккулонодулярная доля) и частично старым мозжечком, входящими в медиальную червячную зону. Получая и обрабатывая импульсацию от вестибулярных рецепторов, от приорецепторов аппарата движения и рецепторов кожи, от зрительных и слуховых рецепторов, мозжечок способен оценить состояние мышц, положение тела в пространстве и через ядра шатра, используя вестибуло-, ретикуло- и руброспинальный тракты, произвести перераспределение мышечного тонуса, изменить позу тела и сохранить равновесие. Нарушение равновесия является наиболее характерным симтомом поражения ар-хицеребеллума.
2. Координация выполняемого движения осуществляется старым и новым мозжечком, входящим в промежуточную (околочервячную) зону. В кору этой части мозжечка поступает импульсация от проприорецепторов, а также импульсация от мо-торной коры большого мозга, представляющая собой программу произвольного движения. Анализируя информацию о программе и выполнении движения (от проприорецепторов), мозжечок способен через свое промежуточное ядро, имеющее выходы на красное ядро и моторную кору, осуществить координацию позы и выполняемого целенаправленного движения в пространстве, а также исправить направление движения. Например, подходя к двери, мы поднимаем руку, чтобы нажать кнопку звонка. Вначале наше движение носит ориентировочный характер; мы так же поднимали бы руку, чтобы поправить прическу, надеть очки. Однако на каком-то этапе это движение становится только движением к кнопке, и, чтобы палец попал именно на кнопку, нужна определенная согласованность действий мышц-антагонистов, причем тем большая, чем ближе цель движения. Внешне движение к цели идет по прямой, без резких изгибов траектории, но эта внешняя «гладкость» движения требует постоянного перераспределения «внимания» центральных регуляторных аппаратов с одной группы мышц на другую. Нарушение координации движения является наиболее характерным симптомом нарушения функции промежуточной зоны мозжечка.
3. Мозжечок участвует в программировании движений, что осуществляется его полушариями. Кора мозжечка получает импульса-цию преимущественно из ассоциативных зон коры большого мозга через ядра моста. Эта информация характеризует замысел движения. В коре нового мозжечка она перерабатывается в программу движения, которая в виде импульсов вновь поступает через таламус в премоторную и моторную ко"ру и из нее через пирамидную и экстрапирамидную системы - к мышцам. Контроль и коррекция более медленных программированных движений осуществляются мозжечком на основе обратной афферентации преимущественно от проприорецепторов, а также от вестибулярных, зрительных, тактильных рецепторов. Коррекция быстрых движений из-за малого времени их выполнения осуществляется путем изменения их программы в самом мозжечке, т.е. на основе обучения и предшествующего опыта. К таким движениям относятся многие спортивные упражнения, печатание на пишущей машинке, игра на музыкальных инструментах.
В. Двигательные функции мозжечка играют важную роль в регуляции мышечного тонуса, сохранении позы, координации выполняемых движений, в программировании планируемых движений. Если мозжечок не выполняет своей регуляторной функции, то у человека наблюдаются расстройства двигательных функций. Эти расстройства проявляются различными симптомами, которые связаны друг с другом.
1. Дистопия (distonia - нарушение тонуса) - повышение или понижение тонуса мышц. При повреждении мозжечка наблюдается повышение тонуса мышц-разгибателей. Характер влияния на тонус мышц определяется частотой генерации импульсов нейронов ядра шатра. При высокой частоте (30-300 имп/с) тонус мышц-разгибателей снижается, при низкой (2-10 имп/с) - увеличивается. В случае повреждения мозжечка активируются нейроны вестибулярных ядер и ретикулярной формации продолговатого мозга, которые активируют мотонейроны спинного мозга. Одновременно активность пирамидных нейронов снижается, а, следовательно, снижается их тормозное влияние на те же мотонейроны спинного мозга. В итоге, получая возбуждающие сигналы от продолговатого мозга при одновременном уменьшении тормозных влияний от коры большого мозга, мотонейроны спинного мозга активируются и вызывают гипертонус мышц-разгибателей.
2. Астения (astenia - слабость) - снижение силы мышечного сокращения, быстрая утомляемость мышц.
3. Астазия (astasia, от греч. а - не, stania - стояние) - утрата способности к длительному сокращению мышц, что затрудняет стояние, сидение.
4. Тремор (tremor - дрожание) - дрожание пальцев рук, кистей, головы в покое; этот тремор усиливается при движении.
5.Дисметрия (dismetria - нарушение меры) - расстройство равномерности движений, выражающееся либо в излишнем, либо в недостаточном движении. Больной пытается взять предмет со стола и проносит руку мнмо предмета (гиперметрия) или не доносит ее до предмета (гипометрия).
6. Атаксия (ataksia, от греч. а - не, 1taksia - порядок) - нарушение координации движений. Здесь ярче всего проявляется невозможность выполнения движений в нужном порядке, в определенной последовательности. Проявлениями атаксии являются также адиадохокинез, асинергия, пьяная - шаткая походка. При адиадохокинезе человек не способен быстро вращать ладони вниз-вверх. При асинергии мышц он не способен сесть из положения, лежа без помощи рук. Пьяная походка характеризуется тем, что человек ходит, широко расставив ноги, шатаясь из стороны в сторону.
7. Дизартрия (disartria- расстройство организации речевой моторики). При повреждении мозжечка речь больного становится растянутой, слова иногда произносятся как бы толчками (скандированная речь).
Данные о том, что повреждение мозжечка ведет к расстройствам движений, которые были приобретены человеком в результате обучения, позволяют сделать вывод, что само обучение идет с участием мозжечковых структур, а следовательно, мозжечокпринимает участие в организации процессов высшей нервной деятельности. При повреждении мозжечка страдают когнитивные процессы.
После операции частичного удаления мозжечка возникают симптомы его повреждения, которые затем исчезают. Если на фоне исчезновения мозжечковых симптомов нарушается функция лобных долей мозга, то мозжечковые симптомы возникают вновь. Следовательно, кора лобных долей большого мозга компенсирует расстройства, вызываемые повреждением мозжечка. Механизм данной компенсации реализуется через лобно-мосто-мозжечковый тракт.
Г. Мозжечок за счет своего влияния на сенсомоторную область коры может изменять уровень тактильной, температурной, зрительной чувствительности.
Удаление мозжечка приводит к ослаблению силы процессов возбуждения и торможения, нарушению баланса между ними, развитию инертности. Выработка двигательных условных рефлексов после удаления мозжечка затрудняется, особенно при формировании локальной, изолированной двигательной реакции. Точно так же замедляется выработка пищевых условных рефлексов, увеличивается скрытый (латентный) период их вызова.
5.7. ПРОМЕЖУТОЧНЫЙ МОЗГ
Промежуточный мозг расположен между средним и конечным мозгом, вокруг III желудочка мозга. Он состоит из таламической области и гипоталамуса. Таламическая область включает в себя таламус, метаталамус (коленчатые тела) и эпиталамус (эпифиз). В литературе, посвященной вопросам физиологии, метаталамус объединяется с таламусом, эпифиз рассматривается в эндокринной системе.
Таламус - парный ядерный комплекс, занимающий преимущественно дорсальную часть промежуточного мозга. В таламусе выделяют до 40 парных ядер, которые в функциональном плане можно разделить на следующие три группы: релейные, ассоциативные и неспецифические. Все ядра таламуса в разной степени обладают тремя общими функциями: переключающей, интегративной и модулирующей.
А. Переключательные ядра таламуса (релейные, специфические) делят на сенсорные и несенсорные.
1. Главной функцией сенсорных ядер является переключение потоков афферентной импульсации в сенсорные зоны коры большого мозга. Наряду с этим происходят перекодирование и обработка информации. Главные сенсорные ядра следующие.
Вентральные задние ядра являются главным реле для переключения соматосенсорной афферентной системы. В них переключаются тактильная, проприоцептивная, вкусовая, висцеральная, частично температурная и болевая чувствительность. В этих ядрах имеется топографическая проекция периферии, поэтому электростимуляция вентральных задних ядер вызывает парастезии (ложные ощущения) в разных частях тела, иногда нарушение «схемы тела» (искаженное восприятие частей тела).
Латеральное коленчатое тело выполняет функции реле для переключения зрительной импульсации в затылочную кору, где она используется для формирования зрительных ощущений. Кроме корковой проекции часть зрительной импульсации направляется в верхние холмики четверохолмия. Эта информация используется для регуляции движения глаз, в зрительном ориентировочном рефлексе.
Медиальное коленчатое тело является реле для переключения слуховой импульсации в височную кору задней части сильвие-вой борозды (извилина Гешля, или поперечная височная извилина).
2. К несенсорным переключательным ядрам таламуса относятся передние и вентральные ядра. Они переключают в кору несенсорную импульсацию, поступающую в таламус из разных отделов головного мозга. В передние вентральное, медиальное и дорсальное ядра импульсация поступает из гипоталамуса. Передние ядра таламуса рассматриваются как часть лимбической системы и иногда обозначаются как «лимбические ядра таламуса».
Вентральные ядра участвуют в регуляции движения, выполняя таким образом моторную функцию. В них переключается нмпуль-сация от базальных ганглиев, зубчатого ядра мозжечка, красного ядра среднего мозга, которая после этого проецируется в моторную и премоторную кору.
Наряду с корковыми проекциями переключательных ядер каждое из них получает нисходящие корковые волокна из той же проекционной зоны, что создает структурную основу для взаиморегулирующих отношений между таламусом и корой
Б. Ассоциативные ядра таламуса включают ядра подушки, медиодорсальное ядро и латеральные ядра. Волокна к этим ядрам приходят не от проводниковых путей анализаторов, а от других ядер таламуса. Эфферентные выходы от этих ядер направляются главным образом в ассоциативные поля коры. В свою очередь кора мозга посылает волокна к ассоциативным ядрам, регулируя их функцию. Главной функцией этих ядер является интегративная функция, которая выражается в объединениидеятельности как таламических ядер, так и различных зон ассоциативной коры полушарий мозга.
Подушка получает главные входы от коленчатых тел и неспецифических ядер таламуса. Эфферентные пути от нее идут в ви-сочно-теменно-затылочные зоны коры, участвующие в гностических (узнавание предметов, явлений), речевых и зрительных функциях (например, в интеграции слова со зрительным образом), а также в восприятии «схемы тела».
В латеральные ядра поступает зрительная и слуховая импульса-ция от коленчатых тел и соматосенсорная импульсация от вентрального ядра. Интегрированная сенсорная информация от этих источников далее проецируется в ассоциативную теменную кору и используется в ее функции гнозиса, праксиса, формировании «схемы тела».
Медиодорсальное ядро получает импульсацию от гипоталамуса, миндалины, гиппокампа, таламических ядер, центрального серого вещества ствола. Проекция этого ядра распространяется на ассоциативную лобную и лимбическую кору. Оно участвует в формировании эмоциональной и поведенческой двигательной активности, а также, возможно, в образовании памяти.
В. Неспецифические ядра составляют эволюционно более древнюю часть таламуса, ее ядра содержат преимущественно мелкие, многоотростчатые нейроны и функционально рассматриваются как производное ретикулярной формации ствола мозга. В неспецифические ядра поступает импульсация от других ядер таламуса по трактам, проводящим преимущественно болевую и температурную чувствительность. В неспецифические ядра непосредственно или через ретикулярную формацию также поступает часть импульсации по коллатералям от всех специфических сенсорных систем. Кроме того, в неспецифические ядра приходит импульсация из моторных центров ствола (красное ядро, черное вещество), ядер мозжечка, от базальных ганглиев и гиппокампа, а также от коры мозга, особенно лобных долей. Неспецифические ядра имеют эфферентные выходы на другие таламические ядра, кору больших полушарий как непосредственно, так и через ретикулярные ядра, а также нисходящие пути к другим структурам ствола мозга, т. е. эти ядра, как и другие отделы ретикулярной формации, оказывают восходящие и нисходящие влияния.
Неспецифические ядра таламуса выступают в роли интегрирующего посредника между стволом мозга и мозжечком, с одной стороны, и новой корой, лимбической системой и базальными ганглиями - с другой, объединяя их в единый функциональный комплекс. На кору мозга неспецифический таламус оказывает преимущественно модулирующее влияние. Разрушение неспецифических ядер не вызывает грубых расстройств эмоций, восприятия, сна и бодрствования, образования условных рефлексов, а нарушает только тонкую регулировку поведения.
Гипоталамус - это вентральная часть промежуточного мозга, макроскопически он включает в себя преоптическую область и область перекреста зрительных нервов, серый бугор и воронку, сосцевидные тела. В гипоталамусе выделяют до 48 парных ядер, которые подразделяются разными авторами на 3-5 групп.
Гипоталамус - многофункциональная система, обладающая широкими регулирующими и интегрирующими влияниями. Однако важнейшие функции гипоталамуса трудно соотнести с его отдельными ядрами. Как правило, отдельно взятое ядро имеет несколько функций. В связи с этим физиология гипоталамуса рассматривается обычно в аспекте функциональной специфики его различных областей и зон. Гипоталамус является важнейшим центром интеграции вегетативных функций, регуляции эндокринной системы, теплового баланса организма, цикла «бодрствование - сон» и других биоритмов; велика его роль в организации поведения (пищевого, полового, агрессивно-оборонительного), направленного на реализацию биологических потребностей, в проявлении мотиваций и эмоций.
БАЗАЛЫ1ЫЕ ГАНГЛИИ
Назальные ганглии расположены в основании больших полушарий и включают три парных образования: бледный шар, филогенетически более позднее образование - полосатое тело и наиболее молодую часть - ограду. Бледный шар состоит из наружного и внутреннего сегментов; полосатое тело включает хвостатое и скорлупу.
А. Функциональные связи базальных ганглиев. Афферентная импульсация в базальные ганглии поступает преимущественно в полосатое тело в основном из трех источников: 1) от всех областей коры непосредственно и через таламус; 2) от черного вещества; 3) от неспецифических ядер таламуса.
Среди эфферентных связей базальных ганглиев можно отметить три выхода:
От полосатого тела пути идут к бледному шару. От бледного шара начинается самый важный эфферентный тракт базальных ганглиев в таламус, в его релейные вентральные ядра, от них возбуждающий путь идет в двигательную кору;
Часть эфферентных волокон из бледного шара и полосатого тела следует к центрам ствола мозга (ретикулярная формация, красное ядро и далее в спинной мозг), а также через нижнюю оливу в мозжечок;
От полосатого тела тормозящие пути идут к черному веществу и после переключения - к ядрам таламуса.
Базальные ганглии являются промежуточным звеном (станцией переключения), связывающим ассоциативную и частично сенсорную кору с двигательной корой. Рассмотрим функции отдельных структур базальных ганглиев.
Б. Функции полосатого тела. 1. Полосатое тело оказывает на бледный шар двоякое влияние - возбуждающее и тормозящее с преобладанием последнего, что осуществляется преимущественно через тонкие тормозные волокна (медиатор ГАМК).
2. Полосатое тело оказывает тормозящее влияние (медиатор ГАМК) на нейроны черного вещества которые в свою очередь оказывают модулирующее влияние (медиатор дофамин) на кортикостриарные каналы связи.
3. Влияние на кору большого мозга: раздражение полосатого тела вызывает синхронизацию ЭЭГ - появление в ней высокоамплитудных ритмов, характерных для фазы медленного сна. Разрушение полосатого тела уменьшает время сна в цикле бодрствование - сон.
4. Стимуляция полосатого тела через хронически вживленные электроды вызывает относительно простые двигательные реакции: поворот головы и туловища в сторону, противоположную раздражению, иногда сгибание конечности на противоположной стороне. Стимуляция некоторых зон полосатого тела вызывает задержку текущей поведенческой деятельности - двигательной, ориентировочной, пищедобывательной. Животное как бы «застывает» в одной позе. При этом на ЭЭГ развиваются медленные высокоамплитудные ритмы. Раздражение некоторых точек полосатого тела приводит к подавлению ощущения боли.
При поражении стриарной системы возникает гипотониче-ски-гиперкинетический синдром, что обусловлено дефицитом тормозящего влияния стриатума на нижележащие двигательные центры, вследствие чего развиваются мышечная гипотония и избыточные непроизвольные движения (гиперкинезы). Гиперкинезы - автоматические чрезмерные движения, в которых участвуют отдельные части тела, конечности. Они возникают непроизвольно, исчезают во сне и усиливаются при произвольных движениях и волнении.
Отдельные виды гиперкинезов связывают с поражением определенных структур стриарной системы. При поражении оральной части полосатого тела возникают насильственные движения в мускулатуре лица и шеи, при поражении средней части - в мускулатуре туловища и рук. Поражение каудальной части полосатого тела вызывает гиперкинезы в ногах. Конкретные симптомы поражения полосатого тела весьма разнообразны.
Атетоз - медленные червеобразные, вычурные движения в дистальных отделах конечностей (в кистях и стопах). Могут наблюдаться в мускулатуре лица: выпячивание губ, перекашивание рта, гримасничанье, прищелкивание языком. Обычно атетоз связывают с поражением крупных клеток стриарной системы. Характерным его признаком является образование преходящих контрактур (зразтиа глоЫН$), которые придают кисти и пальцам своеобразное положение. У детей нередко наблюдается двусторонний, двойной атетоз при подкорковых дегенерациях. Гемиатетоз бывает значительно реже.
Гемибаллизм - размашистые бросковые движения в конечностях, чаще всего в руках в виде взмаха «крыла птицы». Насильственные движения при гемибаллизме производятся с большой силой, их трудно прекратить. Возникновение гемибаллизма связывают с поражением подбугорного ядра (люисово тело), расположенного под зрительным бугром.
Наряду с быстро реагирующей на раздражители первой активизирующей системой, в которую входят проводящие пути, существует еще и неспецифическая система замедленного ответа на внешние импульсы, которая является филогенетически более древней, чем остальные структуры мозга и напоминает диффузный тип нервной системы. Эта структура называется ретикулярной формацией (РФ) и состоит из более чем 100 ядер, соединенных между собой. РФ простирается от ядер таламуса и субталамуса до промежуточной зоны спинного мозга верхних шейных сегментов.
Первые описания РФ были сделаны немецкими морфологами: в 1861 г. К. Рейхертом и в 1863 г. О. Дейтерсом, который и ввел термин РФ; большой вклад в ее изучение внес В.М. Бехтерев.
Нейроны, составляющие РФ разнообразны по величине, строению и функции; имеют обширно разветвленное дендритное дерево и длинные аксоны; их отростки густо переплетаются, напоминая сеть (лат. reticulum - сеточка, formatio - образование).
Свойства ретикулярных нейронов :
1. Мультипликация (умножение импульса) и амплификация (получение конечного большого результата) – осуществляется благодаря сложному переплетению отростков нейронов. Поступающий импульс многократно умножается, что в восходящем направлении дает ощущение даже небольших стимулов, а в нисходящем направлении (ретикулоспинальные пути) позволяет вовлекать в ответную реакцию множество структур НС.
2. Генерирование импульсов . Д. Моруцци доказал, что большинство нейронов РФ постоянно генерирует нервные разряды с частотой порядка 5-10 в секунду. Различные афферентные стимулы суммируются с этой фоновой активностью ретикулярных нейронов, вызывая в одних из них ее учащение, а в других торможение.
3. Полисенсорность . Почти все нейроны РФ способны отвечать на на раздражения самых различных рецепторов. Тем не менее, часть из них реагируют на кожные раздражения и на свет, другие на звуковые и кожные раздражения и т. д. Таким образом, полного смешивания афферентных сигналов в ретикулярных нейронах не происходит; в их связях есть частичная внутренняя дифференциация.
4. Чувствительность к гуморальным факторам и, особенно, к фармпрепаратам. Особенно активными оказываются соединения барбитуровой кислоты, которая даже в небольших концентрациях полностью прекращают активность ретикулярных нейронов, при этом не действуя на спинальные нейроны или на нейроны коры больших полушарий.
В целом же, РФ характеризуется разлитыми рецептивными полями, длительным латентным периодом ответа на периферическую стимуляцию и слабой воспроизводимостью реакции.
Классификация :
Существует топографическая и функциональная класификация РФ.
I. Топографически вся ретикулярная формация может быть подразделена на каудальный и ростральный отделы.
1. Ростральные ядра (ядра среднего мозга и верхней части моста, связанные с промежуточным мозгом) – отвечают за состояние возбуждения, бодрствования, настороженности. Ростральные ядра оказывают локальное влияние на определенные зоны коры больших полушарий. Поражение этого отдела вызывает сонливость.
2. Каудальные ядра (моста и промежуточного мозга, связаны с ядрами черепных нервов и спинным мозгом) – выполняют моторную, рефлекторную и вегетативную функции. Некоторые ядра в процессе эволюции получили специализацию - вазомоторный центр (депрессорные и прессорные зоны), дыхательный центр (экспираторный и инспираторный), рвотный центр. Каудальный отдел РФ оказывает более разлитое, генерализованное влияние на обширные зоны головного мозга. Поражение этого отдела вызывает бессонницу.
Если рассматривать ядра РФ каждого отдела мозга, то РФ таламуса образуют капсулу латерально вокруг зрительных бугров. Они получают импульсы от коры и дорзальных ядер таламуса. Функция ретикулярных ядер таламуса – фильтрация сигналов, идущих через таламус к коре полушарий; проекция их на другие ядра таламуса. В целом, они влияют на всю поступающую сенсорную и когнитивную информацию.
Ядра РФ среднего мозга включают покрышечные ядра: nuclei tegmentalis dorsalis et ventralis , nucleus cuneiformis . Они получают импульсы через fasciculus mammillo-tegmentalis (Gudden) , который является частью мамиллоталямического пути.
РФ моста, образованная околосрединными (парамедианными) ядрами, не имеет четких границ. Эти ядра участвуют в скоординированном движении глаз, фиксированном взгляде и саккадном движении глаз (быстрые синхронные движения глаз). РФ моста лежит кпереди и латерально от медиального продольного пучка, получает импульсы по нервным волокнам от верхних бугорков четверохолмия через предорзальные нервные волокна и от передних зрительных полей посредством лобномостовых связей.
Латеральную РФ образуют преимущественно ядра РФ продолговатого мозга. Эта структура имеет множество ганглиев, вставочных нейронов вокруг черепных нервов, которые служат для модулирования связанных с ними рефлексов и функций.
II. Функционально ядра РФ подразделяются на вертикальные образования:
1. Срединный столб (ядра шва) – узкий парный столб клеток вдоль срединной линии ствола мозга. Простираются от продолговатого до среднего мозга. Дорзальные ядра шва синтезируют серотонин.
2. Медиальный столб (голубоватое пятно) – относится к РФ. Клетки голубоватого пятна синтезируют норэпинефрин, аксоны идут к областям коры, ответственным за возбуждение (бодрствование).
3. Латеральный столб (серое вещество вокруг Сильвиева водопровода) – (входит в состав лимбической системы) – клетки имеют опиоидные рецепторы, что способствует эффекту обезболивания.
Функция РФ:
1. Регуляция сознания путем изменения активности корковых нейронов, участие в цикле сон /бодрствование, возбуждение, внимание, обучаемость - когнитивные функции
2. Обеспечение эмоциональной окраски сенсорным стимулам (ретикулолимбические связи)
3. Участие в жизненно важных вегетативных реакциях (сосудодвигательный, дыхательный, кашлевой, рвотный центры)
4. Реакция на боль – РФ проводит болевые импульсы к коре и образует нисходящие аналгезирующие пути (влияет на спинной мозг, частично блокируя передачу болевых импульсов от спинного мозга к коре)
5. Привыкание – это процесс, в котором мозг учится игнорировать незначительные повторяющиеся стимулы извне, отдавая предпочтение новым стимулам. Пример – способность спать в переполненном шумном транспорте, сохраняя способность проснуться на сигнал автомобиля или крик ребенка
6. Соматодвигательный контроль – обеспечивается ретикулоспинальными путями. Эти пути отвечают за мышечный тонус, равновесие, положение тела в пространстве, особенно при его движении
7. Формирование интегрированных реакций организма на стимулы, например, сочетанная работа речедвигательного аппарата, обшая двигательная активность.
Связи РФ
Аксоны РФ связывают практически все структуры мозга между собой. РФ морфологически и функционально связана со спинным мозгом, мозжечком, лимбической системой и корой больших полушарий головного мозга.
Часть аксонов РФ имеет нисходящее направление и образует ретикулоспинальные пути, а часть восходящее (спиноретикулярные пути). Возможна также циркуляция импульсов по замкнутым нейронным цепям. Таким образом, существует постоянный уровень возбуждения нейронов РФ, вследствие чего обеспечиваются тонус и определённая степень готовности к деятельности различных отделов центральной нервной системы. Степень возбуждения РФ регулируется корой больших полушарий головного мозга.
1. Спиноретикулярные (спиноретикулокортикальные) пути (восходящая активизирующая ретикулярная система) - получают импульсы от аксонов восходящих (сенсорных) путей общей и специальной чувствительности. Соматовисцеральные волокна идут в составе спиноретикулярного тракта (переднебоковой канатик), а также в составе проприоспинальных путей и соответствующих путей от ядра спинального тройничного тракта. К ретикулярной формации приходят также пути от всех других афферентных черепномозговых нервов, т.е. практически от всех органов чувств. Дополнительная афферентация поступает от многих других отделов головного мозга - от моторных областей коры и сенсорных областей коры, мозжечка, базальных ядер, красного ядра, от таламуса и гипоталамуса. Эта часть РФ отвечает за процессы возбуждения, внимания, бодрствования, обеспечивает эмоциональные реакции, важные при познавательном процессе. Поражения, опухоли этой части РФ вызывают снижение уровня сознания, активности психической деятельности, в частности когнитивных функций, двигательной активности, синдром хронической усталости. Возможны сонливость, проявления оглушенности, общей и речевой гипокинезии, акинетического мутизма, сопора, в тяжелых случаях - кома.
2. Ретикулоспинальные пути (нисходящие ретикулярные связи) – могут оказывать как стимулирующее влияние (отвечают за мышечный тонус, вегетативные функции, активирует восходящую РФ), так и угнетающее (способствуют плавности и точности произвольных движений, регулирует мышечный тонус, положение тела в пространстве, вегетативные функции, рефлексы). Обеспечиваются множеством эфферентных связей - нисходящие к спинному мозгу и восходящие через неспецифические таламические ядра к коре головного мозга, гипоталамусу и лимбической системе. Большинство нейронов образует синапсы с двумя - тремя дендритами разного происхождения, такая полисенсорная конвергенция характерна для нейронов ретикулярной формации.
3. Ретикуло-ретикулярные связи .
Сложное строение человеческого мозга открывает тайны нашего поведения, объясняет законы мыслительной деятельности, протекание эмоций и чувств. Каждое полушарие головного мозга отвечает за свои специфические функции и задачи (например, известно, что правое отвечает за логику, а левое - за воображение и фантазию), но имеются также структуры, которые обеспечивают единую и согласованную работу всей центральной нервной системы. Одной из таких структур является ретикулярная формация.
Ретикулярная формация - это участок стволового отдела мозга, представленный разветвленной сетью нервных клеток и ядер, соединяющих между собой различные отделы головного мозга. В отличие от других структур, - например, таламуса, гипоталамуса, мозжечка, - которые имеют некую цельную форму (ядра, железы), ретикулярная формация не представлена единым морфологическим образованием, а является «сетью» (от латинского reticulum — сеть) дендритов и аксонов, которые с разной степенью плотности проникают между отделами и структурами головного мозга, объединяя их между собой и обеспечивая их совместную деятельность.
Метафорически выражаясь: если наш мозг представить в виде некоего изделия, скажем, рубашки, то ретикулярная формация - это нити, которыми рубашка сшита. Ретикулярная формация пронизывает структуры продолговатого, среднего мозга и моста, имеет непосредственные связи с мозжечком, спинным мозгом, таламусом и опосредованные - с вышележащими отделами: гипоталамусом, зрительными ядрами и корой.
В состав ретикулярной формации входит огромное количество нейронов с разветвленными дендритами и длинными аксонами, за счет чего становится возможным передавать нервные импульсы в различные отделы головного и спинного мозга. При этом можно выделить две наиболее крупные группы нейронных скоплений:

Основное предназначение ретикулярной формации состоит в сенсорном анализе многочисленных сигналов, поступающих от различных отделов ГМ.
За счет тесных связей со спинным мозгом она также принимает самое активное участие в моторной регуляции, начиная от глотательного рефлекса и заканчивая сложными двигательными операциями. Кроме того, ретикулярная формация осуществляет активирующее воздействие на весь ГМ в целом, участвуя в регуляции циклов сна и бодрствования.
В общем виде функции ретикулярной формации сводятся к следующим:
Отличительные особенности работы ретикулярной формации связаны, прежде всего, с определенными свойствами ее :
Нервные клетки, входящие в состав ретикулярной формации, обладают повышенной способностью к тоническому возбуждению. Это означает, что бо́льшая часть нейронов находятся в постоянном возбуждении и генерирует нервные импульсы, которые передаются в вышележащие отделы ГМ. Такая тоническая активность обусловлена рядом факторов:
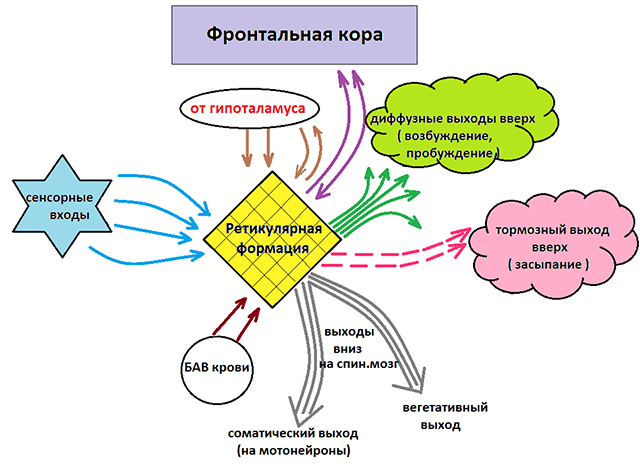
Как уже отмечалось, ретикулярная формация оказывает возбуждающее и тормозящее действие на различные отделы ГМ. При этом можно выделить два отдела, которые специализируются на передаче возбуждения в те или иные структуры мозга.
Нисходящий отдел: представлен вегетативными и двигательными центрами и оказывает нисходящее влияние на отделы спинного мозга. Соответствующие нейронные скопления регулируют деятельность дыхательного, сосудодвигательного, слюноотделительного центров, а также центров, отвечающих за построение простых и сложных двигательных реакций. Это указывает на решающую роль именно центральной нервной системы в регуляции даже элементарных безусловных рефлексов. Стимуляция нисходящего отдела приводит к затормаживанию спинальных центров и вызывает в естественной среде состояние глубоко сна (спать «без задних ног»). Тот же эффект можно вызвать и искусственным путем, например, вводя человека в состояние транса или наркоза.
Восходящий отдел: представлен нервными волокнами, соединяющими структуры ретикулярной формации с вышележащими отделами: таламусом, гипоталамусом, мозжечком и корой. Восходящее влияние оказывает стимулирующее действие на кортикальные структуры и обеспечивает активное состояние сознания. Восходящее влияние не прекращается, даже когда мы спим. Если бы наш мозг мог полностью «отключаться», то каждое пробуждение было бы сродни рождению: кто я? Где я? Как я сюда попал? Однако за счет работы ретикулярных структур у нас сохраняется возможность всегда возвращаться в то исходное состояние сознания, в котором мы находились до момента сна. Кроме того, и во время ночного отдыха у нас остается способность реагировать на некоторые жизненно важные раздражители, т.е. мы, как правило, не спим «мертвым сном» и можем проснуться, если рядом зашевелился и заплакал ребенок, что-то громко упало и т.п.
Ретикулярная формация играет значимую роль в интегративной деятельности всего головного мозга. За счет выполнения функции ведущего проводника всех видов нервных импульсов во все отделы ЦНС, ретикулярная формация находится в постоянной работе. Чрезмерные умственные и эмоциональные перегрузки вредны для мозга в целом и для ретикулярной формации, в частности. К счастью, своевременный прием седативных препаратов может (за счет повышенной восприимчивости нейронов к химическому воздействию) быстро исправить ситуацию и нормализовать состояние.
Однако возможны и менее благоприятные исходы. Повреждение возможно в результате черепно-мозговых травм, онкологических заболеваний головного мозга, инфекционных поражений.
Основным проявлением неблагополучия является потеря сознания.
Нарушение восходящих связей обнаруживает себя в состоянии апатии, слабости, повышенной сонливости, двигательной растороможенности, нарушениях ночного сна. Нередки сопутствующие вегетативные расстройства.
Ретикулярная формация известна с 1845 г., описана Дейтерсом (O.F.C. Deiters) в 1885 г. В настоящее время продолжается ее изучение. Ретикулярная формация располагается между задними и боковыми рогами шейных сегментов спинного мозга, в покрышке мозгового ствола, в центральном ядре зрительного бугра. Представляет собой комплекс анатомически и функционально взаимосвязанных нейронов, окруженных множеством волокон, идущих в различных направлениях к ядерным структурам и проводящим путям (рис. 30).
Рис. 30. : 1 - восходящие пути; 2 - нисходящие пути; 3 - специфический (лемнисковый) чувствительный проводящий путь; 4 - пирамидальный путь.
Ретикулярная формация воспринимает все импульсы (болевые, температурные, световые, звуковые и пр.), однако в ней нет специализированных нейронов. Поэтому одни и те же нейроны воспринимают различные импульсы и передают их в различные отделы мозга, во все участки коры. Ретикулярная формация - это вторая афферентная система головного мозга, его неспецифическая структура . Она имеет двухсторонние связи со всеми структурами головного и спинного мозга (рис. 31, 32).
 Рис. 31. : 1; 2; 3 - специфический (лемнисковый) чувствительный проводящий путь; 4 - коллатерали, соединяющие специфический чувствительный путь с ретикулярной формацией мозгового ствола; 5 - восходящая активирующая система ретикулярной формации; 6 - генерализованное влияние ретикулярной формации на кору головного мозга.
Рис. 31. : 1; 2; 3 - специфический (лемнисковый) чувствительный проводящий путь; 4 - коллатерали, соединяющие специфический чувствительный путь с ретикулярной формацией мозгового ствола; 5 - восходящая активирующая система ретикулярной формации; 6 - генерализованное влияние ретикулярной формации на кору головного мозга.
 Рис. 32. : 1 - чувствительный нерв, на который наносится стимул (болевое раздражение); 2 - спинной мозг; 3 - симпатические нервы; 4 - надпочечник; 5 - каротидный синус; 6 - гипофиз; 7 - ретикулярная формация. Сплошными стрелками обозначены нервные влияния, пунктирными - гормональные влияния, которые через ретикулярную формацию оказывают активирующее воздействие на кору головного мозга.
Рис. 32. : 1 - чувствительный нерв, на который наносится стимул (болевое раздражение); 2 - спинной мозг; 3 - симпатические нервы; 4 - надпочечник; 5 - каротидный синус; 6 - гипофиз; 7 - ретикулярная формация. Сплошными стрелками обозначены нервные влияния, пунктирными - гормональные влияния, которые через ретикулярную формацию оказывают активирующее воздействие на кору головного мозга.
Структурные элементы ретикулярной формации ствола мозга подразделяют на латеральный и медиальный отделы. В латеральном отделе заканчиваются волокна из различных афферентных систем. К рассеянным клеткам и ядрам ретикулярной формации подходят коллатерали от медиальной и латеральной петель, от чувствительных ядер черепных нервов. От нейронов медиального отдела начинаются эфферентные волокна к двигательным ядрам черепных нервов, к мозжечку, к двигательным ядрам передних рогов спинного мозга.
Основные афферентные пути ретикулярной формации: tr. spinoreticularis - от спинного мозга, tr. tegmentothalamicus - от среднего мозга, reticulothalamicus - от продолговатого мозга и моста, tr. thalamocorticalis - ко всем областям и слоям коры полушарий большого мозга. Сетевидная формация активизирует кору большого мозга и мозжечок.
Кора полушарий большого мозга, в свою очередь, посылает по tr. corticoreticularis импульсы в ретикулярную формацию в составе пирамидных путей. Основным эфферентным трактом является tr. reticulospinalis. Этот путь проводит тонические импульсы к гамма-мотонейронам спинного мозга. Ретикулярная формация регулирует моторное звено, обеспечивая координацию движений, синхронность мышечных сокращений, обеспечивает нестандартные движения, рефлекс равновесия, устанавливает антигравитационный тонус мускулатуры, удерживающий тело над землей. Ретикулярная формация перераспределяет мышечный тонус, что в кризисных ситуациях приводит к мобилизации скрытых резервов организма.
Установлена роль голубоватого пятна и ядер шва в регуляции сна и бодрствования. Голубоватое пятно (locus caeruleus ) находится в верхнелатеральной части ромбовидной ямки. Нейроны этого ядра продуцируют норадреналин, активирующий вышележащие отделы мозга. Особенно высока активность нейронов голубоватого пятна во время бодрствования, во время глубокого сна она угасает почти полностью.
Ядра шва (nuclei raphes ) располагаются по срединной линии продолговатого мозга. Нейроциты этих ядер вырабатывают серотонин, который вызывает процессы разлитого торможения и состояние сна.
Ядра ретикулярной формации продолговатого мозга имеют связи с вегетативными ядрами Ⅸ, Ⅹ нервов и симпатическими ядрами спинного мозга. Поэтому они участвуют в регуляции сердечной деятельности, дыхания, тонуса сосудов, секреции желез и так далее.
Ядра Кахаля и Даркшевича, относящиеся к ретикулярной формации среднего мозга, к медиальному продольному пучку (fasciculus longitudinalis medialis ), имеют связи с ядрами третьей, четвертой, шестой, восьмой, девятой, десятой и одиннадцатой парой черепных нервов. Они координируют работу этого пучка, обеспечивая сочетанные повороты головы и глаз при изменении позы или при поиске источника звука, фиксацию взгляда. (Данные движения совершенно необходимы при трудовых и игровых актах).
Эти связи объясняют вегетативные нарушения при вестибулярных перегрузках. Рассеянные нейроны ретикулярной формации выступают в качестве вставочных нейронов охранительных рефлексов глотательного, роговичного (рис. 33), кашлевого рвоты, зевоты, чихания и др.
 Рис. 33. : 1 - рецепторы, расположенные в роговице; 2 - глазная ветвь тройничного нерва; 3 - ложноуниполярная клетка чувствительного узла тройничного нерва; 4 - ассоциативный нейрон - рассеянная клетка ретикулярной формации; 5 - клетка двигательного ядра лицевого нерва; 6 - круговая мышца глаза.
Рис. 33. : 1 - рецепторы, расположенные в роговице; 2 - глазная ветвь тройничного нерва; 3 - ложноуниполярная клетка чувствительного узла тройничного нерва; 4 - ассоциативный нейрон - рассеянная клетка ретикулярной формации; 5 - клетка двигательного ядра лицевого нерва; 6 - круговая мышца глаза.







